The Carian alphabet is a script evolutionary missing link that was conjectured by Revesz [32] to have existed somewhere in western Anatolia as a common ancestor of the Cypriot syllabary and the Old Hungarian alphabet. This situation is illustrated in Fig. 1

Minoan is an Ugric Language In this section we consider the relationship between the Minoan language as recorded in Linear A [14, 41] and Cretan Hieroglyphs [26, 42] and the Uralic language family. The Uralic language family consists of a Finno-Ugric branch and a Samoyedic branch. The Finno-Ugric branch is further divided into a Finno-Permic and an Ugric branch [18]. The Ugric branch is composed of the Hungarian, Khanty and Mansi languages [18]. Linguists have studied the Uralic languages for over two hundred years and identified sets of words that characterize the nodes of this family tree (see Honti [17]). Minoan, the language of Linear A, is an unknown language. Nevertheless, ancient Greek preserves many words from the Minoan language. Beekes [4] collected in a dictionary all the non-Indo-European vocabulary of ancient Greek. While Beekes [4] often identifies the non-Indo-European words of ancient Greek as having unknown origin, we have found corresponding cognate words within the Uralic language family for many of them. We give some examples of these cognate pairs in Tables 7, 8 and 9, which show some apparent cognate ancient Greek and Uralic, Finno-Ugric and Ugric word pairs. In Tables 7, 8, and 9 the similar consonant sounds are highlighted by red, inserted glide consonants are highlighted by blue, and omitted sounds are indicated by underscores. The Hungarian words and their cognates in Tables 7 and 8 are based on the Hungarian etymological dictionary of Zaicz [43]. The Hungarian words and most of their Khanty and Mansi cognates in Table 9 are based on Honti [17]
The ancient Greek words are from the ancient Greek etymological dictionary of Beekes [3, 4]. The associations of the ancient Greek and the Uralic cognates are our work. There were some earlier dictionaries of Greek and Hungarian by J. Aczél in 1926 and more recently by Varga [37], but they completely ignored Finno-Ugric linguistics. Their dictionaries lack any etymological considerations and list words that are not true cognates but medieval or later borrowings. Their dictionaries also contain several false cognates. Nevertheless, they deserve some credit for bringing the issue of larger than expected similarities between the Greek and the Hungarian vocabularies to attention. Tables 7, 8 and 9 have some striking implications. Clearly, the Ugric word cognates are the most remarkable because the Ugric words are unique to the Ugric branch according to Honti [17]. While there are strong Greek and Hungarian connections because Greek missionaries and merchants frequently visited Hungary, there is no similar relationship between Greek and Khanty or Mansi. Hence we have to suppose that Minoan is a previously overlooked Ugric language. The only logical assumption can be that Minoan separated from the Ugric branch and came to Crete before the arrival of proto-Greek speakers sometime around 1450 BC, when the Linear B supplanted the Linear A writing according to the archeological record. The author’s previous decipherments of the Phaistos Disk [30] and Cretan Hieroglyph inscriptions [31] also suggest that the Minoan language was Finno-Ugric. That proposal was received with some skepticism on a geographic ground because it was difficult to imagine how the Minoans could have arrived to Crete from any previously proposed Finno-Ugric homeland. This situation has led us to the consideration of the Hattic language of Anatolia, as described below.

Hattic is an Ugric Language Usually a language family spreads over a connected area. Hence it looks strange that Minoan culture existed primarily in Crete, while Khanty and Mansi live on the eastern side of the Ural Mountains. However, the gap between these two areas can be explained if Minoans migrated to Crete from the north, probably the eastern or northern costal areas of the Black Sea via Anatolia, that is, present day Turkey. If there was such a migration through Turkey, then it also had to occur in very ancient times. According to archeologists in those ancient times, the Hattic culture occupied most of northern and central Turkey [29]. This naturally raises the question whether Hattic is also related to Minoan and whether it could also be an Ugric language. In this section, we consider this issue because if there is a relation between Minoan and Hattic, then Hattic could also help to reconstruct the Minoan language. Linguists generally consider Hattic to be an language isolate. The only exception that we are aware of is that recently, Alexey Kassian [29] suggested some of the following language similarities between Hattic and the Yeniseian languages, Ket and Kott
from
Establishing the West-Ugric Language Family with Minoan, Hattic and Hungarian by a Decipherment of Linear A PETER Z. REVESZ Department of Computer Science and Engineering University of Nebraska-Lincoln Lincoln, NE 68588-0115 USA revesz@cse.unl.edu, cse.unl.edu/~revesz/
www.researchgate.net/publication/333816923_Establishing_the_West-Ugric_Language_Family_with_Minoan_Hattic_and_Hungarian_by_a_Decipherment_of_Linear_A
This old language family might have had some input into Dravidian based on analysis by the same author and some influence on Sumerian. The existence of words in Dravidian probably from LBA due to ocean trade etc.. might have had a massive influence on Dravidian people
WSEAS Transactions on Information Science and Applications
Print ISSN: 1790-0832, E-ISSN: 2224-3402
Volume 16, 2019
Sumerian Contains Dravidian and Uralic Substrates Associated with the Emegir and Emesal Dialects
Author: Peter Z. Revesz
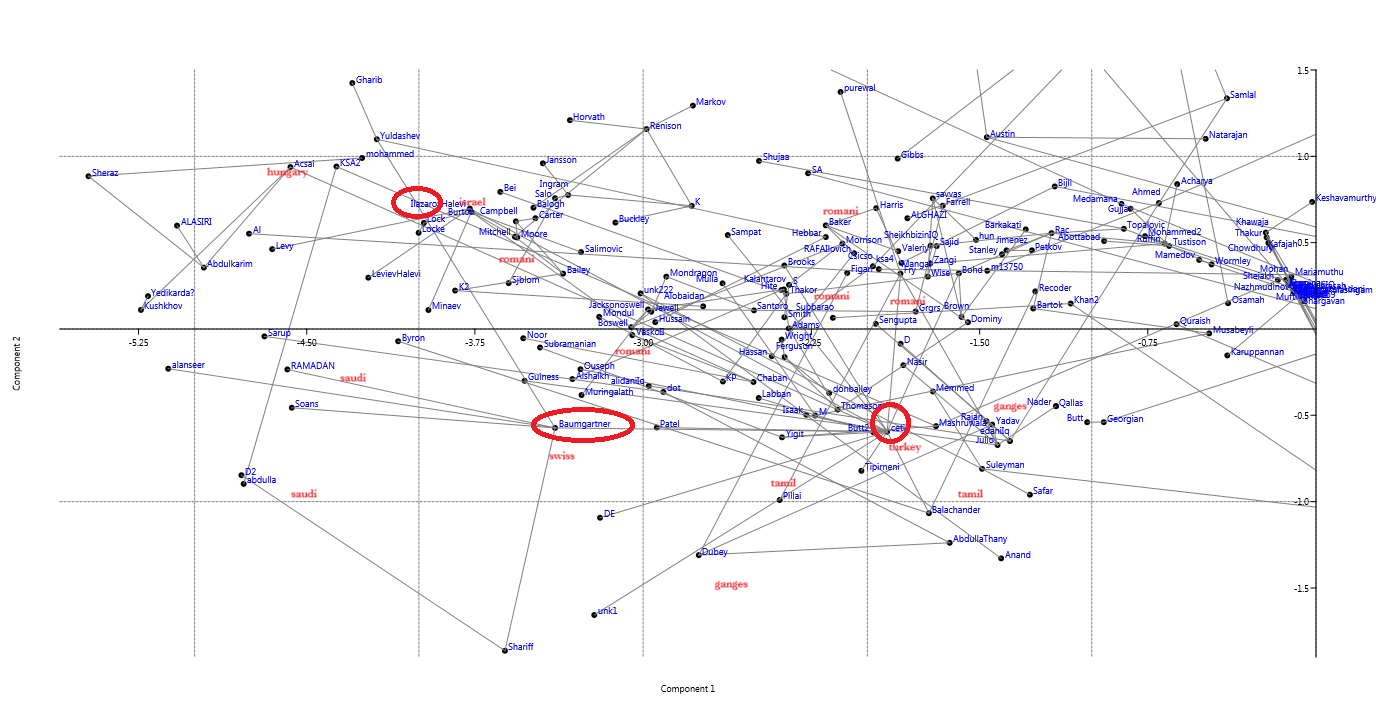
Abstract: Data mining the Sumerian vocabulary reveals a dichotomy of the cognate associations of the Emeĝir and the Emesal dialects, with the former having mostly Dravidian and the later mostly Uralic cognates, indicating that Sumerian arose by the combination of two languages from those language families. The data mining also reveals a distribution pattern of Proto-Uralic, Proto-Finno-Ugric, Proto-Ugric and Proto-Hungarian cognates that indicates that Sumerian is farther than Minoan from Hungarian, although all are West-Ugric.
Dr Peter Z Revesz also published another datamining paper that showed Sumerian is the main language influence into Minoan and later Uralic and Hungarian. Also Dravidian shares some input from these too.
Print ISSN: 1790-0832, E-ISSN: 2224-3402
Volume 16, 2019
Sumerian Contains Dravidian and Uralic Substrates Associated with the Emegir and Emesal Dialects
Author: Peter Z. Revesz
Abstract: Data mining the Sumerian vocabulary reveals a dichotomy of the cognate associations of the Emeĝir and the Emesal dialects, with the former having mostly Dravidian and the later mostly Uralic cognates, indicating that Sumerian arose by the combination of two languages from those language families. The data mining also reveals a distribution pattern of Proto-Uralic, Proto-Finno-Ugric, Proto-Ugric and Proto-Hungarian cognates that indicates that Sumerian is farther than Minoan from Hungarian, although all are West-Ugric.
Dr Peter Z Revesz also published another datamining paper that showed Sumerian is the main language influence into Minoan and later Uralic and Hungarian. Also Dravidian shares some input from these too.
It is massive coincidence since I share the West Ugric lactose persistence and also some ancient Greek/Minoan chunks of DNA not found widely in the present Greeks