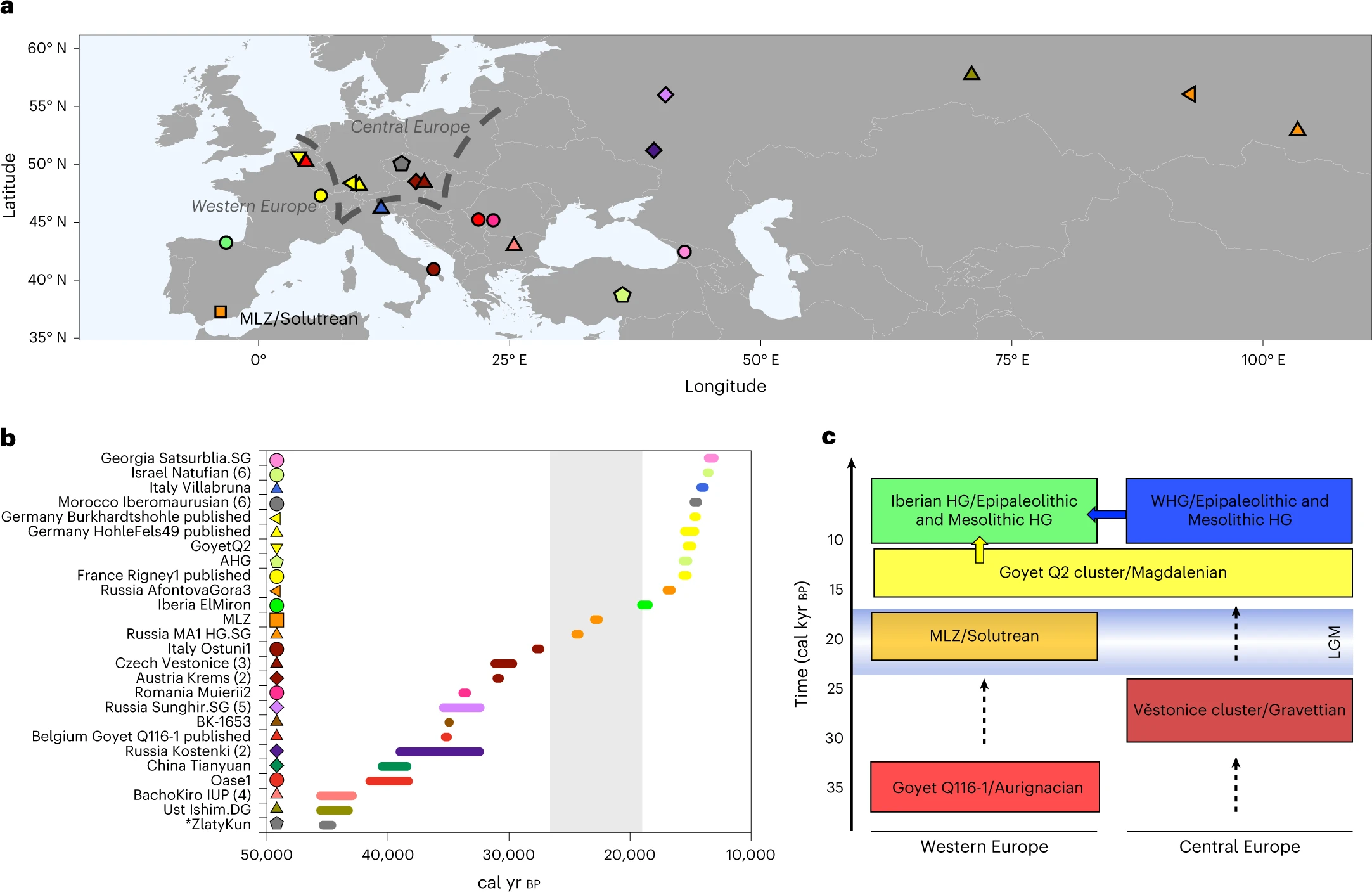
Timeline for AMH evolution in South Asia based on genetic, archaeological, climatological and linguistic evidence. Black and grey portions of the arrow represent Pleistocene and Holocene, respectively. Blue sections correspond to periods of climate changes: dryer periods between 35 and 30 ka, Last Glacial Maximum ~18 ka, Younger Dryas ~12 ka and the " 4.2 ka " event. Lineages in red stand for the putative Late Glacial/postglacial genetic influx from West Eurasia; green for migrations from West Eurasia around the Pleistocene/Holocene transition, orange for the Neolithic period and blue for the genetic events in the last 4 ka
Maternal lineages primarily reflect earlier, pre-Holocene processes, and paternal lineages predominantly episodes within the last 10 ka. In particular, genetic influx from Central Asia in the Bronze Age was strongly male-driven,consistent with the patriarchal, patrilocal and patrilineal social structure attributed to the inferred pastoralist early Indo-European society. This was part of a much wider process of Indo-European expansion, with an ultimate source in the Pontic-Caspian region, which carried closely related Y-chromosome lineages, a smaller fraction of autosomal genome-wide variation and an even smaller fraction of mitogenomes across a vast swathe of Eurasia between 5and 3.5 ka
This result suggests that an ancient western ancestry may have been disguised by further re-expansions ofhaplogroup M in South Asia. Several branches of M(M38, M65, M45, M5b, M5c, M34, M57, M33a) display signals of dispersals from the east and the centre datingt o ~45–35 ka, and M4’67 (which is only separated by asingle mutation from the root of M), with a possible origin in central India, displays an extraordinary multi-branching structure dating to 38.0 [30.1; 46.0] ka, suggesting a major expansion at that time. If we considerthat a root type of M could have survived for ~10,000 yearsafter it arose (as is evident from modern clades within thatage range), it is plausible that re-expansion created a sec-ondary founder effect within M that decreased the overallage estimates. Such a scenario would impact even moreon ρthan ML estimates, which is indeed what we see(Table 1). An expansion 45–35 ka would also fit well with the palaeoenvironmental and archaeological evidence[2, 67, 68], and is further supported by an increment inNeassociated with M across South Asia from ~40 ka(Additional file 1: Figure S1).The next major discernible signal in indigenous lineages begins ~12 ka, at the Pleistocene/Holocene transition.Various star-like clades dating 12–9 ka suggest a rapid ex-pansion across the Subcontinent, namely M6a1a (11.4 ka),M18a (9.2 ka), M30d (12.1 ka), R8b1 (11.6 ka) and U2b2(9.2 ka), all from a southern source; and R30c + 373(12.4 ka), from the west. An increment in Neis also ob-served at this time in the BSP for haplogroup M in thewest and south (Additional file 1: Figure S1).We also see a further increment in the last few millennia.BSPs for M in the west and centre show an increment inthelast2.5ka(Additionalfile1:FigureS1),associated withthe emergence of several subclades in the west (M2a3a +4314, M2a1b, M2c + 1888 + 146, M30a2, M5a3b, M6a1 +5585 + 146 + 1508) and centre (M2a1a1b, M3b, M3a1a,M63, M5a2a2 + 234, M5a3a and M61a + 5294).
West Eurasian mtDNA lineages in South Asia: Multiple dispersals from the northwest since the LGM Prehistoric West Eurasian lineages make up almost 20%of the South Asian genetic pool overall.
The Pakistani Muslim Balochi, Brahui and Makrani
carry ~15% of the Near Eastern/Arabian component
(yellow), which is carried across Europe with the spread
of the Early Neolithic [75, 77]. However, this component
is virtually absent in other South Asians (including
Muslims) except for Jewish groups (supporting previous
mtDNA evidence for little genetic input from Arabia
into Indian Muslim populations [78]
The Pakistani Muslim Balochi, Brahui and Makrani
carry ~15% of the Near Eastern/Arabian component
(yellow), which is carried across Europe with the spread
of the Early Neolithic [75, 77]. However, this component
is virtually absent in other South Asians (including
Muslims) except for Jewish groups (supporting previous
mtDNA evidence for little genetic input from Arabia
into Indian Muslim populations [78]
from A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals
Pedro Soares et all
Subcontinent points to heavily
sex-biased dispersals